
Efficacité des harpes électriques pour réduire la pression de prédation de Vespa velutina et conséquences sur le développement des colonies d’abeilles mellifères
Sandra V Rojas-Nossa,Damian Dasilva-Martins,Salustiano Mato,Caroline Bartolomé,Xulio Maside,Josefina Garrido
Première publication:18 août 2022
https://doi.org/10.1002/ps.7132
Résumé
CONTEXTE
Vespa velutina est devenue une espèce préoccupante dans les régions envahies d’Europe et d’Asie, en raison de ses impacts sur la biodiversité, l’apiculture et la société. Ce frelon, féroce chasseur d’insectes pollinisateurs, constitue une menace sérieuse pour la biodiversité et les services de pollinisation. Malgré les efforts en cours, son extermination en Europe continentale est entravée par un manque de méthodes de contrôle efficaces, de sorte que des mesures d’atténuation efficaces sont les principales préoccupations. Les objectifs de ce travail étaient : (i) d’étudier les effets de la prédation de V. velutina sur les colonies d’abeilles mellifères, et (ii) d’évaluer l’efficacité des harpes électriques pour réduire la pression de chasse et la prédation. Nous avons évalué la pression de prédation et comparé les performances des colonies d’abeilles mellifères, le poids corporel des ouvrières et la survie hivernale pour les abeilles protégées par rapport auxcolonies non protégées dans 36 ruches expérimentales réparties sur trois ruchers.
RÉSULTATS
Les harpes électriques protégeaient les abeilles mellifères en réduisant la pression de prédation et en atténuant ainsi la paralysie de recherche de nourriture. Par conséquent, l’activité de recherche de nourriture, le revenu pollinique, la production de couvain et le poids corporel des ouvrières étaient plus élevés dans les colonies protégées qui, à leur tour, présentaient une plus grande survie hivernale que celles qui n’étaient pas protégées, en particulier sur les sites présentant des niveaux de prédation intermédiaires à élevés.
CONCLUSION
La prédation de V. velutina affecte l’activité de recherche de nourriture, la reproduction, le poids corporel et la survie des colonies d’ Apis mellifera . Les harpes électriques contribuent significativement à atténuer l’impact de ce frelon envahissant sur les ruchers ; cependant, ils devraient être déployés en tandem avec des mesures supplémentaires pour préserver les stocks de colonies d’abeilles mellifères, comme faciliter l’accès aux sources de nourriture pour les colonies pendant les périodes de pression de prédation la plus élevée. © 2022 Les auteurs. Pest Management Science publié par John Wiley & Sons Ltd au nom de la Society of Chemical Industry.
1. INTRODUCTION
La croissance démographique humaine nécessite l’expansion des surfaces cultivées, et donc une plus forte demande de services de pollinisation. 1 Cependant, les populations d’insectes sont en déclin dans le monde entier, 2 en raison de multiples facteurs de stress en interaction, notamment la perte d’habitat, le manque de ressources florales, la propagation de parasites et de maladies et les interactions antagonistes avec les espèces envahissantes. 3 – 5
Vespa velutina Lepeletier 1836, est une espèce envahissante préoccupante dans plusieurs régions européennes et asiatiques en raison de ses impacts sur la biodiversité, l’économie et la santé humaine. 6 , 7 Cet insecte eusocial a un système d’accouplement polyandre et un cycle annuel, 8 qui augmentent leur capacité à élever des populations entières à partir de la propagule minimale : une seule fondatrice accouplée. 9
Bien qu’originaire d’Asie de l’Est, V. velutina a été accidentellement introduit en Corée du Sud 10 et en France. 11 Il s’est étendu avec succès dans les zones voisines touchant plus de dix nouveaux pays au cours de la dernière décennie, 12 , 13 et continue de se propager.
Vespa velutina a un régime généraliste, se nourrissant de nectar floral, de sève, de fruits, de miellat et de miel comme sources de glucides, tandis que les protéines sont obtenues à partir de la chasse aux arthropodes et des charognards, 8 entraînant des impacts sur la biodiversité par la prédation et la compétition pour les ressources. 14 – 17 Notamment, V. velutina chasse les pollinisateurs dans les parcelles de fleurs, entraînant une modification de l’abondance et des habitudes de recherche de nourriture, avec des effets néfastes sur la pollinisation des plantes indigènes. 18 C’est aussi un chasseur efficace et persistant d’abeilles mellifères à l’entrée des ruches, provoquant des pertes de colonies et menaçant ainsi la durabilité de l’apiculture dans les zones envahies. 19
Les abeilles mellifères occidentales ( Apis mellifera Linnaeus, 1758) adoptent des comportements défensifs en réponse aux attaques de frelons. 20 Néanmoins, ceux-ci sont inefficaces pour réduire la pression de la chasse, permettant aux frelons de précéder intensivement les abeilles à l’entrée des ruches. 21 Cela augmente l’expression des gènes associés au stress oxydatif, ce qui peut nuire à la santé des abeilles dans les colonies attaquées. 22 En outre, la chasse aux frelons déclenche un phénomène connu sous le nom de paralysie de recherche de nourriture, c’est-à-dire l’arrêt de l’activité de recherche de nourriture dans les colonies prédatées, ce qui entraîne à son tour une réduction du stockage du miel. 23 – 25La modélisation de la population indique que ce phénomène est la cause de l’augmentation de la mortalité hivernale observée dans les ruchers des zones envahies, 23 cependant aucune preuve empirique confirmant une telle relation n’a encore été rapportée.
Afin de réduire l’impact des frelons sur les colonies d’abeilles, un certain nombre de pratiques ont été mises en place dans les ruchers, mais plusieurs d’entre elles ont une faible efficacité et/ou représentent une pression supplémentaire sur la biodiversité, et d’autres doivent encore être développées. 26 Les museaux de ruche sont une méthode respectueuse de la biodiversité qui réduit la paralysie de la recherche de nourriture et améliore probablement la survie des colonies, 25 , 27 mais le dispositif ne capture ni ne tue les frelons, son impact est donc local. Les harpes électriques sont une autre méthode couramment utilisée dans les ruchers, avec un coût économique moyen et de faibles effets secondaires sur la faune indigène. 26Ils se composent d’un cadre avec des fils électrifiés parallèles verticaux qui produisent un choc électrique lorsqu’ils sont touchés par un insecte volant. Sa performance réelle dans la réduction de l’activité de chasse aux frelons dans les colonies d’abeilles mellifères nécessite une évaluation plus approfondie, mais des observations préliminaires suggèrent que cela pourrait être une méthode efficace et sélective. 28
Dans une zone insulaire, l’éradication de V. velutina a été possible grâce à une combinaison d’outils efficaces, à un engagement coordonné de tous les secteurs concernés et à une action opportune. 29 Le contrôle de l’espèce une fois qu’elle est établie est cependant difficile et nécessite le développement et l’évaluation de nouvelles méthodes pour réduire sa propagation et ses effets sur la biodiversité. 30 Ici, nous avons surveillé les paramètres de la colonie dans 36 ruches expérimentales, réparties sur trois ruchers dans des environnements différents. Nos objectifs étaient de (i) étudier les effets de V. velutinaprédation des colonies d’abeilles mellifères en termes d’activité de butinage, de revenu pollinique, de stockage des ressources, de croissance démographique (superficie couverte par le couvain et les adultes), de poids des abeilles et de survie hivernale, et (ii) d’évaluer l’efficacité des harpes électriques pour réduire la pression de chasse et prédation.
2. MATÉRIELS ET MÉTHODES
2.1 Sites d’étude
L’expérience a été réalisée de juillet à octobre 2020 dans trois ruchers de la région sud-ouest de Pontevedra, Galice, Espagne (Informations complémentaires, Fig. S1 ). Pour cette année, une densité de 1,92 nids/km 2 dans la province de Pontevedra et de 0,96 nids/km 2 pour l’ensemble de la communauté autonome de Galice a été signalée aux autorités (Xunta de Galicia, données non publiées). Cependant, il a été suggéré que la densité réelle des nids dans cette région peut être estimée à 14–17 nids/km 2 . 5 Les trois sites présentent des conditions environnementales différentes et chacun représente un écosystème principal de cette région (Fig. S1 ).
2.2 Conception expérimentale
Au total, 36 ruches Langstroth ont été placées dans trois ruchers (12 ruches par rucher). Pour assurer l’homogénéité de l’état et de la génétique des abeilles mellifères, les colonies d’abeilles mellifères occidentales ibériques ( A. mellifera iberiensis Engel, 1999) ont été acquises auprès du même fournisseur avec des reines élevées tôt dans la saison et maintenues dans un rucher sous la même apiculture. pratiques jusqu’à peu de temps avant le transport vers les ruchers expérimentaux.
Au sein de chaque rucher deux traitements ont été mis en place : six ruches protégées de la prédation de V. velutina avec des harpes et six ruches non protégées où V. velutina chassait les abeilles mellifères ad libitum . Les deux groupes de ruches ont été placés dans des coins opposés du rucher avec une distance > 20 m entre eux. Chaque groupe de ruches a été placé en ligne, avec les entrées orientées dans la même direction, avec une séparation de 20 à 30 cm entre elles, et placé sur des bases à 30–40 cm de hauteur.
Les harpes électriques étaient situées perpendiculairement aux entrées des ruches, entre deux ruches contiguës (Fig. 1(a) ). Ces appareils ont été placés dans les ruchers 1 mois avant les premières observations. Au cours de la première semaine, les harpes ont été éteintes pour permettre aux abeilles de s’habituer à leur présence. Le piège est constitué d’un cadre avec des fils verticaux parallèles connectés alternativement aux pôles positif et négatif d’un circuit électrique. 26 Le modèle utilisé ici a une modification pour minimiser les prises accessoires (Fig. S2 ). Les frelons volants reçoivent un choc électrique chaque fois qu’ils touchent deux fils consécutifs (Fig. 1 (b)), les paralysant pendant quelques secondes et tombant dans une cage en dessous avec des parois grillagées qui permettent aux insectes plus petits de s’échapper. Puis les frelons rampent à l’intérieur de la cage jusqu’à tomber dans une bouteille collectrice.

Figure 1
Système de protection : six ruches alignées, séparées de 20 à 30 cm chacune, protégées par cinq harpes électriques (a). Lors de la chasse aux abeilles devant les ruches, les frelons volent entre les fils électrifiés qui les paralysent (b). Ensuite, ils tombent dans une cage d’où s’échappent de plus petits insectes tandis que les frelons tombent dans une bouteille de collecte.
2.3 Captures de V. velutina et d’insectes non ciblés dans les harpes
Des échantillons mensuels d’insectes capturés ont été obtenus. Les insectes piégés ont été identifiés et comptés in situ au niveau de l’ordre, de la famille et, dans le cas des hyménoptères, au niveau de l’espèce. Les captures d’espèces ou de groupes ont été exprimées comme le taux d’insectes capturés par piège et par jour en divisant le nombre d’insectes capturés par le nombre de jours de capture à chaque piège. La sélectivité a été calculée comme la fraction de frelons piégés sur le nombre total d’insectes capturés.
2.4 Pression de prédation de V. velutina sur les ruches
Deux méthodes ont été utilisées pour quantifier la pression de prédation de V. velutina sur les abeilles mellifères de juillet à octobre. La première consistait à compter visuellement le nombre de V. velutina en chassesur une période de 10 min, une fois par mois, entre 10h00 et 11h00, le jour même où les performances des colonies expérimentales ont été évaluées. Un à trois observateurs étaient situés à 3-5 m devant chaque groupe de six ruches expérimentales et comptaient le nombre de frelons arrivés à chaque groupe. Aucune différence entre les comptages n’a été détectée lorsque plusieurs observateurs étaient présents. Pour calculer le taux de chasse par ruche (HRH), le nombre total de frelons observés a été divisé par le nombre de ruches puis exprimé en nombre de frelons/ruche/10 min. Au cours de la même période d’observation, le nombre d’événements de chasse réussis a été quantifié. Le taux de prédation a été estimé comme le nombre d’abeilles mellifères capturées/nombre de frelons chassant en 10 min. La deuxième méthode pour quantifier la présence de V. velutinadevant, les ruches utilisaient des enregistrements vidéo pour calculer l’indice de prédation à l’entrée de la ruche (HEPIX). Une caméra (GoPro Hero7) a été placée à 0,5 m devant chaque ruche pour une durée d’enregistrement de 10 min. Les enregistrements ont été effectués de 11h00 à 13h00. Les vidéos ont été analysées visuellement en laboratoire à l’aide du Behavioral Observation Research Interactive Software (BORIS). 31Les 2 premières minutes ont été ignorées pour éviter toute interférence de l’opérateur dans le comportement des abeilles et pour permettre l’accoutumance à la présence de la caméra. Le nombre optimal de minutes à analyser, pour une efficacité maximale avec une variance minimale, a été fixé à cinq. Chaque minute a été visualisée trois fois par un seul observateur. Une fois pour compter le nombre d’abeilles entrant dans la ruche, une autre pour compter celles qui sortent et une dernière pour enregistrer la présence de V. velutina devant la ruche. HEPIX mesure le temps avec au moins un V. velutina de chasse présent dans l’image vidéo (Informations complémentaires, Tableau S1). Pendant les périodes d’observation, l’observateur ne pouvait pas visualiser la présence ou l’absence des harpes. HRH et HEPIX étaient positivement corrélés (coefficient de corrélation de Pearson ρ = 0,57, df = 142, P < 0,001), cependant, en septembre et octobre à Gondomar, les frelons ont passé moins de temps à chasser près de l’entrée des ruches, mais ont été observés en train de se nourrir à une plus grande distance (Fig. S3(a) ). Pour cette raison, HRH a été utilisé comme variable explicative pour les analyses statistiques.
2.5 Performances des colonies expérimentales d’abeilles mellifères
2.5.1 Activité de recherche de nourriture
Les vidéos ont également été utilisées pour quantifier le nombre d’abeilles butineuses sortant et entrant dans chaque ruche pendant 5 min. Pour ce faire les vidéos ont été visualisées au ralenti (vitesse 0.5) avec le logiciel QuickTime Player.
2.5.2 Revenu pollinique
Un piège à pollen a été installé à l’entrée de chaque ruche (voir Fig. 1(a) ). Une fois par mois, la porte de la trappe à pollen était fermée pour recueillir le pollen apporté par les abeilles ouvrières pendant 24 h. Au laboratoire, le poids du pollen frais a été mesuré avec une balance de précision OHAUS Pioneer® 0,01 mg.
2.5.3 Stockage des ressources et évaluation des individus d’âges différents
Une fois par mois, des cadres ont été extraits des ruches pour évaluer la surface couverte par les rayons remplis de miel ouvert, de miel coiffé et de pollen, et le couvain (œufs, larves et pupes) en utilisant une grille de 5 cm × 5 cm. Pour calculer le nombre d’abeilles mellifères adultes, un comptage visuel des individus a été effectué dans une grille de 10 cm × 10 cm pour chaque côté des cadres, puis estimé pour l’ensemble du cadre et enfin additionné pour calculer le nombre total d’ouvrières au sein de la ruche. . 32 Les mesures ont été effectuées de 10h00 à 16h00 par temps ensoleillé.
2.6 Poids des abeilles
En octobre 2020, 30 abeilles ouvrières ont été collectées à l’intérieur de chaque ruche expérimentale en les brossant d’un ou, au maximum, de deux rayons à couvain. Ceux-ci ont été stockés dans de l’éthanol à 80 % à 4 °C. Au laboratoire, ils ont été séchés dans une étuve à 65 °C pendant 12 h, puis pesés individuellement avec une balance de précision OHAUS Pioneer® 0,01 mg. Le temps de séchage a été optimisé et fixé après réalisation d’une courbe de séchage en mesurant les poids consécutifs de 30 individus toutes les 5 min.
2.7 Survie hivernale
En mars 2021, les ruches ont été inspectées pour évaluer la survie de chaque colonie. Les colonies dépeuplées (avec moins de 50 individus) ou sans reine (avec des ouvrières pondeuses donc sans couvain femelle et avec moins de 50 ouvrières) ont été considérées comme des colonies mortes. Le nombre minimum d’individus a été fixé en dessous du niveau critique de survie de la colonie selon le modèle de la ruche sociale pour une colonie régulière. 33Pour comparer la survie hivernale des colonies sous différentes pressions de prédation, un numéro a été attribué à chaque ruche en fonction de sa survie (1 = vivant, 0 = mort). La prédation globale a été évaluée comme une variable semi-quantitative : nulle (HRH = 0), faible (HRH < 1), intermédiaire (1 < HRH 0,5 < 1,5) et élevée (HRH > 1,5). Pour affecter chaque groupe de ruches à l’une de ces catégories, les HRH devaient se situer dans ces seuils depuis plus de 3 mois (Fig. S3 ).
2.8 Analyses statistiques
Une corrélation de Pearson a été utilisée pour étudier la relation entre HRH et HEPIX. Un test de somme des rangs de Wilcoxon a été utilisé pour comparer les différences entre le taux de capture de V. velutina dans les harpes et la capture de taxons non ciblés. Pour tester l’effet de la période d’échantillonnage sur le taux de capture de V. velutina et la sélectivité des harpes, des modèles mixtes linéaires généralisés (GLMM) gonflés à zéro avec des distributions binomiales négatives ont été ajustés à l’aide de la fonction d’optimisation non linéaire par défaut glmmTMB. Ces modèles incluaient le mois comme effet fixe et le site comme effet aléatoire. Voir tableau S2pour une description complète des modèles et des scripts. Pour tester les différences de HRH entre les traitements (ruches protégées avec des harpes versus non protégées) et la période d’échantillonnage, un GLMM, avec une distribution binomiale négative, a été ajusté à l’aide de la fonction d’optimisation non linéaire par défaut du glmmTMB. Le modèle incluait le traitement et le mois comme effets fixes et le site comme effet aléatoire. Pour tester l’effet de HRH sur la quantité de couvain, d’adultes, de butineuses, de pollen apporté par les butineuses (en milligrammes) et sur les réserves de miel et de pollen des ruches expérimentales au cours de la période d’échantillonnage, des GLMM avec des distributions binomiales négatives ont été ajustées à l’aide du glmmTMB par défaut non- fonction d’optimisation linéaire. Les modèles incluaient HRH et le mois comme effets fixes etsite comme effet aléatoire. L’absence de surdispersion a été testée via des tests de dispersion basés sur la simulation (tableau S2 ), avec le package « DHARMa » pour R. 34 Une correction séquentielle de Bonferroni a été appliquée pour corriger les tests de comparaison multiple. Le poids des ouvrières des colonies protégées versus non protégées a été comparé avec un modèle linéaire généralisé (GLM) utilisant une distribution gaussienne. Le modèle incluait le traitement , le site et leur interaction en tant qu’effets fixes. Pour analyser la survie des colonies, un GLM avec une distribution binomiale a été ajusté. La prédation globale et le poids moyen des travailleurset leur interaction ont été inclus comme effets fixes. Les analyses ont été réalisées à l’aide du logiciel RStudio version 1.2.5033. Les packages ‘glmmTMB’ et ‘car’ ont été utilisés pour le montage des GLMM. 35 , 36
3. RÉSULTATS
3.1 Captures de V. velutina et d’insectes non ciblés dans les harpes
Vespa velutina représentait> 90% des 4359 insectes capturés dans les harpes électriques (tableau S3 ). Le taux de capture de V. velutina variait dans le temps ( χ 2 = 78,88 ; df = 5, P < 0,01 ; voir Fig. 2 ). Dans l’ensemble, les captures de V. velutina étaient significativement plus élevées que les captures de taxons non ciblés [moyenne ± écart type (SD) = 1,6 ± 2,0 individus/piège/jour et moyenne ± SD = 0,17 ± 0,2 individus/piège/jour respectivement ; W = 1889, df = 90, P < 0,01], qui se composait principalement d’hyménoptères, comme A. mellifera, suivis des diptères des familles Sarcophagidae, Tachiniidae et Calliphoridae (Tableau S3 ). La sélectivité des harpes était élevée (moyenne ± SD = 0,75 ± 0,34) et est restée élevée dans le temps ( χ 2 = 7,80 ; df = 5, P = 0,168).
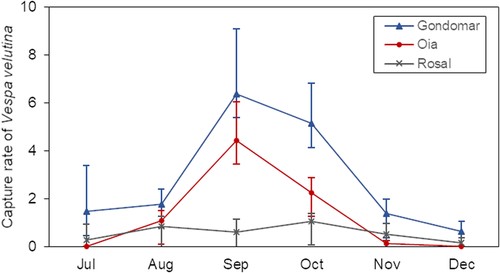
Figure 2
Captures de Vespa velutina par des harpes électriques dans trois ruchers. Les lignes colorées représentent la tendance du taux de capture moyen par site ( n = 5 pour chaque point). Les barres verticales représentent les écarts types.
3.2 Pression de chasse et taux de prédation
Dans l’ensemble, les ruches protégées ont montré une pression de prédation inférieure de 88,8 % à celle des ruches non protégées (Fig. 3 ). Le taux de frelons chassant devant les ruches (HRH) variait significativement entre les traitements (protégés avec des harpes versus non protégés) et à travers les mois d’échantillonnage ( χ 2 = 42,6, df = 1, P < 0,001 et χ 2 = 24,76, df = 3 , P < 0,001 respectivement). L’activité de chasse des frelons a montré une tendance générale à augmenter au cours de l’étude (Fig. S3(a) ). HRH était la plus élevée à Gondomar (moyenne ± SD = 1,4 ± 1,41), moyenne à Oia (moyenne ± SD = 0,6 ± 0,58) et la plus faible à O Rosal (moyenne ± SD = 0,2 ± 0,28). Le succès des frelons chassant les abeilles (Fig. S3(b)) avait des tendances similaires à HRH tout au long de la saison (Fig. S3 (a) ), sauf pour le site à forte pression de prédation où le HRH le plus élevé ne coïncidait pas avec le taux de prédation le plus élevé.
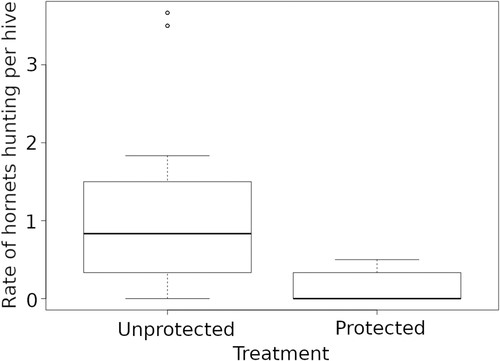
figure 3
Pression de prédation de Vespa velutina sur les ruches protégées et non protégées. Les diagrammes à moustaches représentent les médianes (lignes horizontales en gras), les quartiles (boîtes), les centiles 2,5 à 97,5 (lignes verticales) et les valeurs aberrantes (points vides).
3.3 Performances des colonies expérimentales d’abeilles mellifères
3.3.1 Activité de recherche de nourriture
Le nombre de travailleurs à la recherche de nourriture a été affecté par la pression de chasse et le mois (tableau 1 ). La paralysie de recherche de nourriture enregistrée dans les ruches non protégées sur le site avec la plus forte prédation (Gondomar) a commencé en août et a persisté pendant toute l’étude malgré la diminution de la prédation observée en septembre (Figs 4(a) et S3 (a) ). La paralysie de recherche de nourriture s’est produite principalement dans les colonies d’abeilles mellifères qui ont subi des taux de chasse supérieurs à 0,8 frelons/ruche/10 min (d’après les données des figures 4 (a) et S3 (a) ). Dans le site avec la HRH la plus élevée, les colonies protégées avaient des tailles de population plus importantes dans la ruche que celles protégées en août et septembre (Fig. 4 (b) ), précisément lorsque ces colonies ont montré une paralysie (Fig. 4(a) ). Nous avons observé des ouvrières de tous âges restant à l’intérieur de la ruche dans des colonies paralysées. Outre la recherche de nourriture, d’autres comportements courants tels que la collecte de résines pour la production de propolis ou les activités hygiéniques, telles que le nettoyage des vols, l’enlèvement des personnes malades ou des cadavres, ont également cessé.
Tableau 1. Effets de la pression de chasse, du site et de la période de l’année sur les performances de la colonie d’abeilles mellifères
| Variables de réponse | Prédicteurs | x 2 | P |
| Revenu de pollen (mg) | SAR | 33,72 | <0,001 |
| Mois | 84,25 | <0,001 | |
| Miel non coiffé | SAR | 0,03 | 0,854 |
| Mois | 27,59 | <0,001 | |
| Miel coiffé | SAR | 0,31 | 0,580 |
| Mois | 4.16 | 0,245 | |
| Magasins de pollen | SAR | 0,27 | 0,599 |
| Mois | 1,89 | 0,596 | |
| Nichée | SAR | 8h30 | 0,028 |
| Mois | 76,99 | <0,001 | |
| Adultes | SAR | 5.16 | 0,138 |
| Mois | 150.13 | <0,001 | |
| Butineuses | SAR | 8,85 | 0,024 |
| Mois | 67.27 | <0,001 |
- Le couvain comprend les œufs, les larves et le couvain coiffé. HRH, taux de chasse par ruche. Les résultats significatifs ( P< 0,05), en appliquant la correction séquentielle de Bonferroni, compte tenu des 14 tests effectués, sont indiqués en gras.
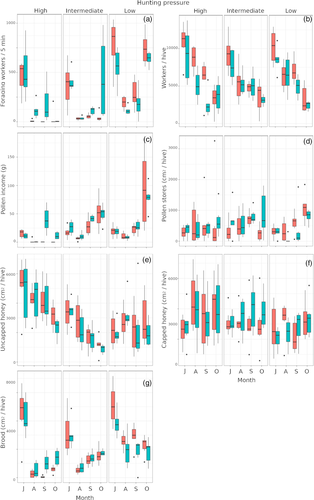
Figure 4
Performance des colonies d’abeilles mellifères et stockage des ressources pour les colonies protégées (barres bleues) et non protégées (barres rouges) dans trois ruchers tout au long de la saison de surveillance.
3.3.2 Ressources
Les ruches non protégées avaient un revenu de pollen plus faible avec un effet significatif du mois (Tableau 1 et Fig. 4(c) ). La quantité de pollen apportée par les butineuses et l’activité des butineuses ont suivi des tendances similaires (Tableau 1 et Fig. 4(c) ). Les réserves de pollen et de miel n’ont pas été significativement affectées par la prédation de V. velutina (tableau 1 et fig. 4 (d, f)).
3.3.3 Performances de la colonie
La quantité de couvain et l’activité de recherche de nourriture ont été significativement affectées par la pression de prédation et le mois (tableau 1 ). La quantité de couvain avait des niveaux plus faibles dans les ruches non protégées que dans les ruches protégées dans le rucher avec la pression de chasse la plus élevée (Fig. 4(g) ).
3.4 Poids des abeilles mellifères
Les ouvrières des ruches non protégées étaient 6,7 % plus légères que celles des ruches protégées. Le GLM a révélé que le traitement, le site et l’interaction traitement X site influençaient le poids des abeilles ( χ 2 = 14,20, df = 1, P < 0,001 ; χ 2 = 95,77, df = 2, P < 0,001 ; χ 2 = 23,19, df = 1, P < 0,001 respectivement).
3.5 Survie
La survie hivernale était de 77,8 % pour les colonies protégées et de 55,6 % pour les colonies non protégées. La survie des colonies non protégées était plus faible sur les sites à pression de chasse intermédiaire et élevée (Fig. 5 ). La survie des colonies d’abeilles mellifères était liée à la pression de chasse globale et à l’interaction entre la pression de chasse et le poids des abeilles, mais pas au poids des abeilles seul ( χ 2 = 10,23, df = 3, P = 0,017 ; χ 2 = 11,43 , df = 3, P = 0,01 ; et χ 2 = 0,02, df = 1, P = 0,90 respectivement).
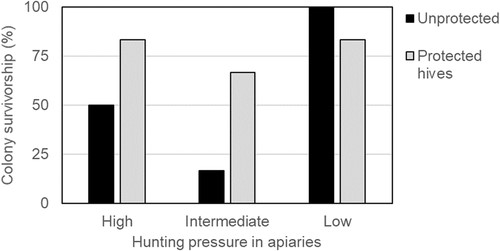
Figure 5
Survie hivernale de colonies expérimentales d’abeilles mellifères (non protégées et protégées avec des harpes électriques dans trois ruchers. n = 6 pour chaque barre).
4. DISCUSSION
Dans cette étude, nous avons examiné l’efficacité des harpes électriques dans la protection des colonies d’abeilles mellifères contre la prédation de V. velutina et avons démontré que cette méthode réduit considérablement la pression de chasse devant les ruches. En comparant les colonies protégées et non protégées, nous avons également détecté des différences dans le comportement de recherche de nourriture, le revenu des ressources, la quantité de couvain et d’adultes et le poids corporel des ouvrières. En raison de la pression de la chasse, les abeilles mellifères entrent en hiver avec une réserve de nourriture réduite, une population réduite et des individus mal nourris. Ainsi, la survie hivernale des colonies est compromise, ce qui entraîne des taux de mortalité hivernale plus élevés pour les colonies non protégées.
4.1 Efficacité et sélectivité des harpes
Les harpes électriques, placées perpendiculairement à une ligne de ruches, sont utiles pour réduire la pression de prédation de V. velutina dans les ruches d’abeilles. Le piégeage des ouvriers de chasse représente un avantage par rapport aux autres méthodes de contrôle, telles que les museaux de ruche ou les pièges appâtés, 25 , 37 – 39 car il réduit efficacement la paralysie de recherche de nourriture de manière ciblée, tout en minimisant les prises accessoires. L’effet protecteur des harpes aide à réduire la paralysie de la recherche de nourriture et permet aux abeilles d’afficher des comportements naturels, y compris la recherche de nourriture, l’hygiène ou l’éventail, nécessaires à la survie et au maintien de la santé des colonies.
Les taux de chasse variaient dans le temps et dans l’espace. Une étude précédente a révélé une augmentation de la pression de prédation au fil du temps, expliquée par la croissance de la population de V. velutina et le besoin de protéines pour élever de nouveaux individus. 21 D’après la corrélation négative entre l’altitude et le nombre de nids observée dans une précédente étude, 40nous nous attendions à trouver un gradient de pression de prédation des frelons reflétant les conditions altitudinales. De manière constante, le rucher situé à la plus haute altitude (Oia), a montré des niveaux de prédation intermédiaires par rapport à celui de Gondomar, situé à moyenne altitude. Cependant, contrairement aux attentes, le rucher situé à l’altitude la plus basse (O Rosal) a présenté le taux de prédation le plus faible. Cela va dans le sens de Monceau et Thiéry, 37 qui ont signalé que la répartition de cette espèce envahissante peut être assez variable dans le temps et dans l’espace, et pourrait être fortement influencée par des processus naturels et/ou anthropiques.
Les GLMM ont révélé qu’en plus de la prédation, des facteurs temporels jouaient également un rôle important sur la quantité de ressources disponibles dans la ruche et l’aptitude de la colonie. La réduction du revenu pollinique a coïncidé avec la paralysie alimentaire sur les colonies non protégées et était probablement liée à la faible quantité de couvain observée au cours des derniers mois de l’étude (Fig. 4 ) . Ces résultats soutiennent l’idée de la paralysie de recherche de nourriture comme principale raison des processus d’affaiblissement et de l’effondrement ultérieur qui affectent les colonies sous l’attaque des frelons. 24Nos preuves suggèrent que dans les ruchers à faible taux de prédation (moins de 1 frelon/ruche/10 min), les abeilles mellifères étaient mieux à même de faire face au prédateur et l’utilisation de harpes électriques était utile pour réduire la prédation à des niveaux négligeables. Cependant, sur les sites où la saison de floraison est plus courte et l’hiver est plus long et avec des températures plus basses (comme à Oia, le site le plus élevé de notre étude), des taux intermédiaires de chasse conduisent à une survie plus faible des colonies dans les ruches protégées et surtout dans les ruches non protégées ( figure 5 ).
Le poids corporel inférieur des ouvrières dans les colonies non protégées fournit une nouvelle preuve du stress physiologique que les abeilles domestiques subissent sous cette nouvelle menace. Le pollen est une ressource d’importance capitale pour les abeilles car c’est une source importante de protéines, de lipides, de vitamines et d’autres nutriments, nécessaires à la production de la gelée royale, avec laquelle les larves sont nourries. 41 , 42 En outre, la qualité et la diversité du pollen influencent la santé et la durée de vie des abeilles mellifères et la qualité du pollen est connue pour affecter le poids des larves chez les abeilles mellifères et les bourdons. 43 – 45Ainsi, une pénurie de revenus en nutriments lors de l’élevage des larves est un mécanisme plausible derrière les travailleurs plus légers. De plus, la plus faible quantité de couvain observée dans les ruches non protégées à la fin de l’étude suggère que le nombre d’ouvrières à hiverner est plus faible que dans les colonies protégées. Ainsi, nos résultats ont révélé qu’outre le faible stockage des ressources alimentaires, deux facteurs sont à l’origine de la mortalité hivernale des colonies d’abeilles mellifères non protégées : une carence du niveau nutritionnel des adultes hivernants et un nombre réduit d’ouvrières.
4.2 Gestion des ruchers en zone envahie
Le seuil des niveaux de prédation nécessaires pour produire des changements significatifs dans les performances et la survie des colonies est inférieur à celui signalé précédemment. 24 Cela peut s’expliquer par plusieurs facteurs non mutuellement exclusifs qui méritent une attention pour les études futures et la gestion des ruchers. L’un d’eux est lié aux facteurs environnementaux qui déterminent la facilité (ou la difficulté) pour les abeilles d’accéder aux ressources pendant la saison de prédation et les conditions d’hivernage.
De plus, nous fournissons des preuves que l’état physiologique des ouvrières des abeilles mellifères est affecté par la prédation par les frelons. Par conséquent, nous encourageons les apiculteurs à faciliter l’accès des abeilles aux diverses ressources florales et d’eau douce, contribuant ainsi à diminuer leur stress physiologique et leur échec de retour à l’extérieur de la ruche. Pour cette raison, lorsque les colonies souffrent de paralysie de recherche de nourriture, le complément d’une alimentation diversifiée riche en tous les nutriments nécessaires est recommandé. Cela doit être fourni non seulement pendant l’hiver pour éviter la famine des colonies faibles mais aussi en automne lorsque les ouvrières qui vont endurer l’hiver sont au stade larvaire.
L’installation de harpes électriques représente un investissement économique initial important pour les apiculteurs qui dépend du nombre de harpes et de son prix commercial. Ceci, à son tour, est lié au nombre et à l’emplacement des ruches. Il a été suggéré un rationnel d’une harpe toutes les deux ou trois ruches, ce qui est probablement inabordable pour les grands ruchers. 26 Aussi, en termes de temps nécessaire au maintien d’un système fonctionnel. En outre, les ruchers doivent souvent être adaptés, et il est suggéré de former des lignes avec une distance réduite entre les ruches afin de réduire la distance entre les entrées de la ruche et la harpe, pour obtenir un effet protecteur plus élevé. 28 Néanmoins, dans cette étude, nous avons observé que dans les ruchers placés dans des sites à forte abondance de V. velutina, avec une ligne compacte de ruches (20 à 30 cm de distance entre elles) et une harpe entre deux ruches consécutives, la réduction de la pression de chasse était significative mais encore insuffisante pour atteindre une prédation nulle. Par conséquent, dans les zones fortement envahies, cette méthode de contrôle devrait être déployée en tandem avec des mesures supplémentaires, telles que la détection et la destruction des nids de V. velutina aux abords des ruchers afin de réduire le nombre de chasseurs et leurs conséquences néfastes sur le miel. performance et survie des colonies d’abeilles.
5. CONCLUSIONS
La pression de prédation exercée par le frelon envahissant V. velutina interfère avec l’activité normale de la ruche et affecte les caractéristiques biologiques des abeilles mellifères, telles que l’activité de recherche de nourriture, la reproduction, le poids corporel et la survie de la colonie. Un nombre réduit de frelons chassant les abeilles devant les ruches pendant une période prolongée suffit à déclencher la paralysie de la colonie. La forte mortalité des colonies d’abeilles mellifères est un phénomène mondial enregistré depuis la fin du siècle dernier. Ici, nous démontrons que les techniques de gestion apicole, telles que l’utilisation de harpes électriques, aident à améliorer la santé des colonies d’abeilles mellifères et à prolonger leur durée de vie dans les zones envahies en réduisant les effets néfastes de V. velutina sur les pollinisateurs . La pression de prédation de V. velutinas’ajoute aux facteurs de stress auxquels sont confrontés les pollinisateurs dans les zones envahies. Il est donc indispensable d’éviter l’implantation de prédateurs efficaces des abeilles, comme le frelon V. velutina ou d’autres espèces envahissantes du genre Vespa , dans les régions à risque. 46 , 47 Ainsi, l’élaboration de plans de prévention et de gestion à long terme est nécessaire, permettant ainsi à l’apiculture de rester viable, tout en préservant la pollinisation des plantes et la biodiversité.
REMERCIEMENTS
Ce travail a été financé par le Programme Interreg Espace Atlantique (Fonds Européen de Développement Régional – FEDER, Union Européenne) : EAPA_800/2018 – Atlantic-Positive. Financement des frais d’accès libre : Universidade de Vigo/CISUG. Des suggestions sur la conception de l’expérience ont été aimablement faites par X. Asorey. Les auteurs remercient D. Boisits, A. Lagoa et Apicultura Niño do Corvo pour leur soutien sur le terrain. TA O’shea-Wheller et P. Kennedy ont contribué à améliorer cet article.
CONFLIT D’INTÉRÊT
Les auteurs déclarent n’avoir aucun intérêt financier lié au travail soumis pour publication. SVRN a été co-auteur du modèle d’utilité ES 1203011-U.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Laisser Un Commentaire